Submitted on: 2025, 29th January; revised on: 2025, 18th April; accepted on: 2025, 8th June
Contributing to the breeding phenology of a temperate nocturnal gecko
20200 Bastia France
Email: dmighe@gmail.com
Abstract. This short descriptive note updates and summarises knowledge on the breeding phenology of the nocturnal Gekkota Euleptes europaea. A forty-year dataset covering the entire range of the species is provided. Between 1982 and 2024, a total of 39 populations were studied in 85 sampling sessions. Reproduction is seasonal, with gravid females appearing from early spring until mid-June when oviposition begins. Hatching occurs from mid-August to early October.
Keywords. Ectotherm, Euleptes europaea, gravid, hatchling, Sphaerodactylidae, squamate.
Understanding the natural history of species, particularly the timing of their life cycles, is critical in a warming world (Parmesan et al., 1999; Ibáñez et al., 2010; Forrest, 2016). Because temperate ecosystems are highly seasonal and reproduction is costly, animals are expected to invest in reproduction when both abiotic and biotic conditions are optimal (Brown and Shine, 2006). In squamates, developmental success is strongly related to temperature and, especially in oviparous species, to moisture. In temperate latitudes, temperature and relative hygrometry vary seasonally. As stated by Van Dyke (2015), the reproductive success of squamates is maximised when reproduction is initiated at the right time of the year. Seasonal variations in temperature and precipitation provide direct cues to the environmental conditions favourable to reproduction, while photoperiod provides an indirect metric of season and should be correlated, at least in part, with seasonal changes in temperature and humidity. Accordingly, squamates appear to use all three as seasonal cues to reproduce (Van Dyke, 2015). The thermal constraints are particularly critical for strictly nocturnal ectotherms living in temperate regions, i.e., in thermal deficit for most of their annual activity period (Saint-Girons and Saint-Girons, 1956; Chukwuka, Monks, and Cree, 2023). Among the Gekkota, all kinds of reproductive strategies have been observed, even in sympatry, ranging from the continuous to the highly cyclical (Vitt, 1986; Righi, Nascimento, and Galdino, 2012)
The aim of this note is to assess whether the reproduction of a nocturnal gecko, Euleptes europaea, is seasonal, as it would be expected since females of most temperate reptile species reproduce during the warmest part of the year (Pianka and Vitt, 2003). However, given the efficient thermoregulatory abilities of this gecko (Delaugerre, 1984) and its dessication-resistant hard-shelled eggs (Mourgue, 1910), one might expect reproductive activity to be less dependent on temperature and humidity, and spread over weeks or months, but still seasonal rather than continuous (Brown and Shine, 2006).
E. europaea is a strictly nocturnal, small-sized, Mediterranean gecko mainly living on islands. Along with suitable climatic conditions, the availability and the quality of retreat sites and egg laying sites (narrow rock cracks) are likely the key factor governing its presence and its abundance. It is no longer considered a pure rock specialist, as recent studies have shown it to be arboreal as well (Salvi et al., 2023; Deso et al., 2024). For comprehensive information on the natural history and the distribution of the study species, see Salvidio, Lanza, and Delaugerre (2010).
The current state of knowledge on the phenology of this species is based on Knoepffler (1973) studies carried out on the island of Port-Cros (Provence) in the 60s and on Delaugerre (1981) studies on Port-Cros in 1978 and 1979. On Port-Cros, mating takes place in March. Gravid females are observed from April to the end of July. Egg-laying begins in mid-May and continues until the end of June or July (two successive clutches). Hatching occurs 65-80 days after oviposition from late July to early October, a female usually lays a pair of eggs, hatchlings measure about 17 mm in Snout-Vent-Lenght (SVL).
In this study, we use a 40-year dataset based on the survey of populations across most of the species range, from Tunisia in the south to Provence in the north-west (from 37.3° to 43° north latitude), including the islands of Sardinia and Corsica (Table 1). The populations of Tuscany and Liguria were not part of the survey. Between 1982 and 2024, a total of 39 populations were studied in 85 sampling sessions (repeated sessions of some populations). Most of the populations studied are Mediterranean landbridge islets, except two from the continental island of Corsica and one from the ‘large’ island of Port-Cros. All of these localities are below 200 m above sea level, and 75% of them are below 50 m. All of the islets are uninhabited and often included in protected areas where access is prohibited. Although they are not subject to direct anthropogenic disturbance, they are affected by the effects of globalisation, such as climate change, rising sea level and colonisation by alien species. Observations were carried out from early April to mid-October. Sampling sessions lasted from 1 to 6 nights per population. Active geckos were searched for using battery-powered lamps, starting one or two hours after dusk and continuing until dawn if necessary. Geckos were carefully caught by hand and temporarily stored in bags. Animals were gently held between thumb and forefinger under the anterior members until their posture was in extension, and SVL was measured to the nearest mm using a digital caliper.
| Region | Population | Geographic coordinates | Date | Day/Month |
|---|---|---|---|---|
| Corsica | Gargalu | 42°22’12”N 8°32’22”E | 2-3/04/1985 | 3/4 |
| Corsica | Toro G | 41°30’33”N 9°22’53”E | 14/04/2005 | 14/4 |
| Provence | Port-Cros | 43°00’18”N 6°23’49”E | 23-26/04/2001 | 23/4 |
| Corsica | Locca | 41°53’09’N 08°36’03’E | 30/04/2010 | 30/4 |
| Corsica | Cornuta | 41°37’55’N 09°22’07’E | 26/05/2017 | 26/5 |
| Provence | Gabinière | 42°59’18”N 6°23’43”E | 25/05/2016 | 25/5 |
| Tunisia | Gallo | 37°33’28”N 8°57’21”E | 3-6/05/2008 | 3/5 |
| Corsica | Gargalu | 42°22’12”N 8°32’22”E | 18-19/05/2022 | 18/5 |
| Corsica | Lavezzu | 41°20’25’N 09°15’15’E | 6-9/05/2024 | 6/5 |
| Corsica | Falaises de Bonifacio | 41°22’37”N 9°10’34”E | 22/05/2020 | 22/5 |
| Corsica | Mezzumare | 41°52’38’N 08°35’34’E | 28/05/2024 | 28/5 |
| Corsica | Nord Pinarellu | 41°40’22”N 9°23’40”E | 25/05/2017 | 25/5 |
| Sardegna | Piana Tavolara | 40°53’18”N 9°39’04”E | 03/05/2022 | 6/5 |
| Sardegna | Porco | 41°10’27”N 9°27’39”E | 29/05/2012 | 3/5 |
| Corsica | Porro | 41°53’20’N 08°36’22’E | 29/05/2014 | 29/5 |
| Sardegna | Spargiotto | 41°15’00”N 9°19’25”E | 08/05/2014 | 29/5 |
| Provence | St Féréol | 43°30’22”N 7°03’28”E | 20/05/2016 | 8/5 |
| Provence | Tradelière | 43°30’56”N 7°04’23”E | 21/05/2016 | 20/5 |
| Corsica | Scandula | 42°22’06”N 8°34’19”E | 06/05/1982 | 21/5 |
| Corsica | Lavezzu | 41°20’25’N 09°15’15’E | 2-4/06/2021 | 2/6 |
| Corsica | Lavezzu | 41°20’25’N 09°15’15’E | 2-4/06/2010 | 2/6 |
| Sardegna | Piana Tavolara | 40°53’18”N 9°39’04”E | 04/06/2024 | 4/6 |
| Sardegna | Reulino Tavolara | 40°52’39”N 9°40’17”E | 04/06/2024 | 4/6 |
| Corsica | Sperdutu G | 41°22’12’N 09°18’18’E | 15/06/2011 | 15/6 |
| Corsica | A Botte | 41°56’32”N 8°35’22”E | 21-28/06/2011 | 28/6 |
| Corsica | Brocciu | 42°38’46”N 8°55’56”E | 29/06/2012 | 29/6 |
| Corsica | Cala d’Alga | 41°52’56’N 08°35’55’E | 26/06/2017 | 26/6 |
| Corsica | Porro | 41°53’20’N 08°36’22’E | 20/06/2011 | 20/6 |
| Corsica | Gargalu | 42°22’12”N 8°32’22”E | 28-29/06/2022 | 28/6 |
| Corsica | Giraglia | 43°01’30’N 09°24’24’E | 21/06/2020 | 21/6 |
| Corsica | Lavezzu | 41°20’25’N 09°15’15’E | 14-17/06/2011 | 14/6 |
| Corsica | Lavezzu | 41°20’25’N 09°15’15’E | 14-15/06/2012 | 14/6 |
| Sardegna | Piana Tavolara | 40°53’18”N 9°39’04”E | 14/06/2022 | 14/6 |
| Provence | Rascas | 43°00’52”N 6°23’21”E | 17-18/06/2008 | 17/6 |
| Sardegna | Reulino Tavolara | 40°52’39”N 9°40’17”E | 13/06/2022 | 13/6 |
| Corsica | Sperdutu g | 41°22’12’N 09°18’18’E | 15/06/2011 | 15/6 |
| Sardegna | Toro | 38°51’40”N 8°24’34”E | 28/06/2015 | 28/6 |
| Corsica | Giraglia | 43°01’30’N 09°24’24’E | 07/07/2014 | 7/7 |
| Corsica | Vacca | 41°33’23’N 09°23’14’E | 09/07/2012 | 9/7 |
| Sardegna | Reulino Tavolara | 40°52’39”N 9°40’17”E | 10/07/2024 | 10/7 |
| Sardegna | Piana Tavolara | 40°53’18”N 9°39’04”E | 12/07/2024 | 12/7 |
| Corsica | A Botte | 41°56’32”N 8°35’22”E | 15/07/2023 | 15/7 |
| Corsica | Gargalu | 42°22’12”N 8°32’22”E | 16;28/07/1990 | 16/7 |
| Corsica | Garganellu | 42°21’55’N 08°32’23’E | 17/07/1986 | 17/7 |
| Corsica | Lavezzu | 41°20’25’N 09°15’15’E | 29/06-4/07/2020 | 29/6 |
| Sardegna | Molarotto | 40°52’27”N 9°46’42”E | 11/07/2024 | 11/7 |
| Corsica | Palazzinu | 42°22’47”N 8°33’01”E | 19-20/07/1985 | 19/7 |
| Corsica | Porragia G | 41°23’30’N 09°15’47’E | 25/07/2024 | 25/7 |
| Corsica | Porri | 42°23’10’N 08°34’55’E | 23-24/07/1983 | 23/7 |
| Corsica | Porri | 42°23’10’N 08°34’55’E | 21/07/1986 | 21/7 |
| Corsica | Toro G | 41°30’33”N 9°22’53”E | 24/07/2014 | 24/7 |
| Corsica | Giraglia | 43°01’30’N 09°24’24’E | 02/08/2022 | 2/8 |
| Corsica | Giraglia | 43°01’30’N 09°24’24’E | 07/08/2023 | 7/8 |
| Corsica | A Botte | 41°56’32”N 8°35’22”E | 11/08/2022 | 11/8 |
| Corsica | 2nd rock Toro P | 41°30’30”N 9°22’47”E | 05/08/1986 | 5/8 |
| Tunisia | Fauchelle | 37°29’39”N 8°52’43”E | 30/08/2022 | 30/8 |
| Corsica | Gargalu | 42°22’12”N 8°32’22”E | 10-11/08/2023 | 10/8 |
| Corsica | Garganellu | 42°21’55’N 08°32’23’E | 12/08/2023 | 12/8 |
| Corsica | Giraglia | 43°01’30’N 09°24’24’E | 5;9/08/2012 | 5/8 |
| Corsica | Lavezzu | 41°20’25’N 09°15’15’E | 02/08/1986 | 2/8 |
| Corsica | Mezzumare | 41°52’38’N 08°35’34’E | 16/08/2012 | 16/8 |
| Corsica | Palazzu | 42°22’49”N 8°32’47”E | 22/08/1986 | 22/8 |
| Corsica | Porragia G | 41°23’30’N 09°15’47’E | 06/08/1985 | 6/8 |
| Corsica | Porragia P | 41°23’35’N 09°15’52’E | 01/08/1986 | 1/8 |
| Corsica | Ruscana | 41°41’00’N 09°24’07’E | 07/08/1986 | 7/8 |
| Corsica | Sperdutu G | 41°22’12’N 09°18’18’E | 04/08/1986 | 4/8 |
| Corsica | Toro G | 41°30’33”N 9°22’53” | 04/08/1986 | 4/8 |
| Corsica | Vacca | 41°33’23’N 09°23’14’E | 2-4/08/1985 | 2/8 |
| Corsica | Giraglia | 43°01’30’N 09°24’24’E | 29/08/2015 | 29/8 |
| Corsica | A Botte | 41°56’32”N 8°35’22”E | 3-6/09/2010 | 3/9 |
| Sardegna | Carpa | 41°17’15”N 9°21’45”E | 06/09/2011 | 6/9 |
| Corsica | Giraglia | 43°01’30’N 09°24’24’E | 12/09/2000 | 12/9 |
| Corsica | Ruscana | 41°41’00’N 09°24’07’E | 10-13/09/2012 | 10/9 |
| Tunisia | Gallina | 37°33’07”N 8°56’56”E | 19/09/2024 | 19/9 |
| Corsica | Gargalu | 42°22’12”N 8°32’22”E | 28/09/2023 | 28/9 |
| Provence | Rascas | 43°00’52”N 6°23’21”E | 02/09/1985 | 2/9 |
| Provence | Gabinière | 42°59’18”N 6°23’43”E | 13/10/2003 | 13/10 |
| Corsica | Gargalu | 42°22’12”N 8°32’22”E | 04/10/2022 | 4/10 |
| Corsica | Giraglia | 43°01’30’N 09°24’24’E | 04/10/2014 | 4/10 |
| Corsica | Giraglia | 43°01’30’N 09°24’24’E | 06/10/2012 | 6/10 |
| Corsica | Garganellu | 42°21’55’N 08°32’23’E | 14/10/2024 | 14/10 |
| Corsica | Sperdutu g | 41°22’12’N 09°18’18’E | 1-2/10/1984 | 1/10 |
| Corsica | Garganellu | 42°21’55’N 08°32’23’E | 01/10/2022 | 11/10 |
| Provence | Gabinière | 42°59’18”N 6°23’43”E | 13/10/2010 | 13/10 |
| Corsica | Ruscana | 41°41’00’N 09°24’07’E | 7-11/10/2008 | 7/10 |
Females were checked for gravidity by visual examination of the belly, the eggs -even smaller than 2 mm- being visible through the thin skin (Figure 1). After examination, geckos were later released in the area of the original sighting. In order to avoid double-counting, when more than one night of prospection was planned, the animals were marked with non-toxic acrylic ink before being released. Gravidity was examined in 36 populations (68 sessions); censuses without females and those without examination of females for eggs were discarded [N = 2450 geckos, of which 729 adult females, of which 262 with visible eggs (Table S1)]. Limitation: It is possible that, just after the first egg-laying, very small eggs in formation may have gone unnoticed by some females.

The presence of newborn juveniles (Figure S1) with a SVL < 22 mm, i.e., aged less than 4-6 weeks (Delaugerre, 1981), was recorded together with the number of juveniles and subadults [N = 2276 geckos; 318 subadults, 487 juveniles of which 127 < 22 mm SVL ] Populations without juvenile observations were discarded (Table S2). Limitation: at hatching, the newborns measure ≈ 17 mm SVL (weight ≈ 0.25 g). They grow rapidly in the first month (+ 4.5 mm) (Delaugerre, 1981; 2003). For this reason, we set the threshold at > 22 mm SVL, below which the young were considered to be only a few weeks old. Even if stature is highly variable from one microinsular population to another (Delaugerre and Cheylan, 1992; Delaugerre et al., 2019; Delaugerre and Corti, 2020), with tendencies towards gigantism and - rarely - dwarfism, we believe that these variations have little effect on these early stages of growth, as hatching size appears to be invariable between populations (M. Delaugerre pers. obs.). This statement might not be true for the dwarf population of Gargalu island.
So, to sum up; the two landmarks of reproduction in this study are: 1) observing the timing of egg-bearing females; the frequency of non-pregnant females marking the egg-laying period, and 2) measuring hatchlings in natural populations; the presence of newborns marking the hatching period.
Gravid females were observed from early April to early October. 258 females out of 439 (59%) were gravid between 2 April and 25 July, while 4 females out of 290 (1.4%) were gravid between 1 August and 14 October (Table S1, Figure 2).
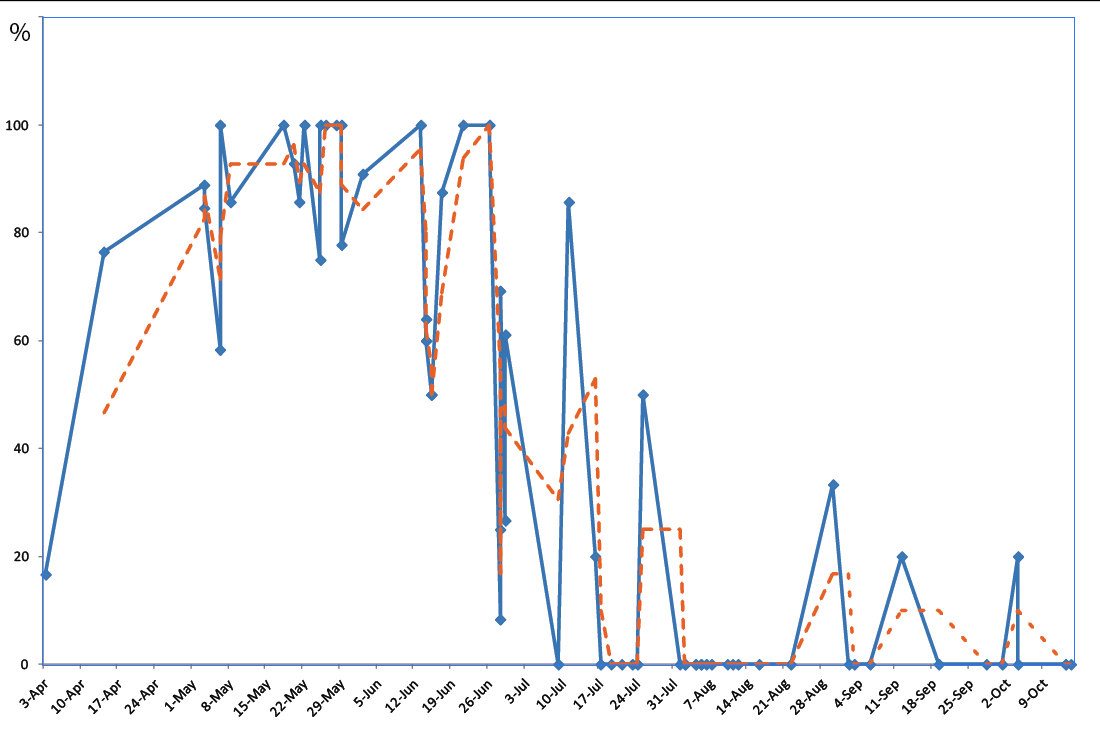
All young age classes (sub-adults, juveniles and hatchlings under 22 mm SVL) were observed from early April to mid-October. From early April to the end of July, the juvenile class represented 18.7% of the population and hatchlings represented 9.9% of the juveniles, but only 3% if the Gargalu population is excluded from the analysis (N geckos 978, N juveniles 183, N hatchlings 18, N = 5 without Gargalu). From early August to mid-October, juveniles account for 23.7% of the population and hatchlings for 35.5% of the juveniles (N geckos 1298, N juveniles 307, N hatchlings 109); see Table S2 and Figure 3.
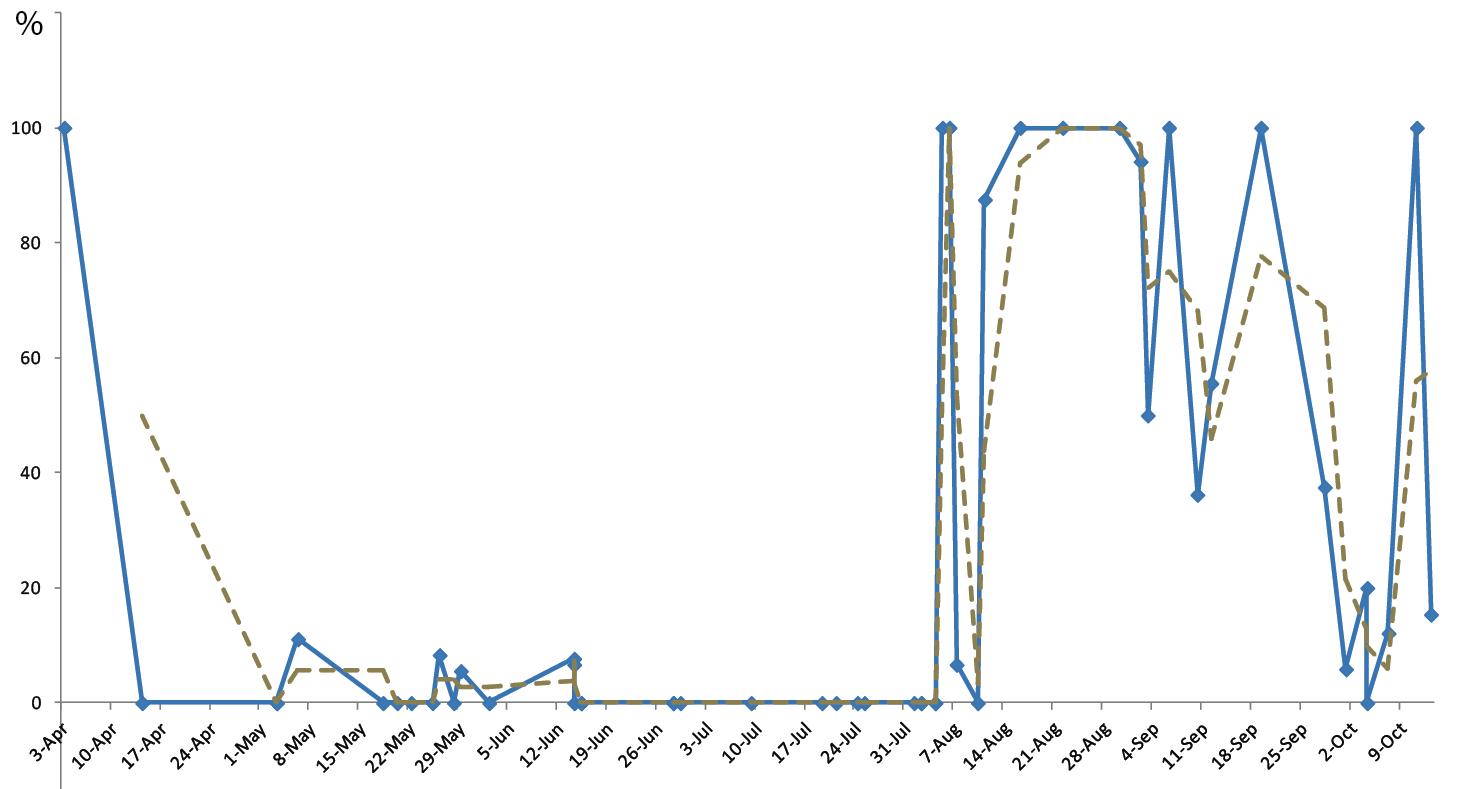
As this species is able to live above 1000-1500 m above sea level (Salvidio, Lanza, and Delaugerre, 2010), our results only apply to the phenology of the coastal populations.
At sea level, the European leaf-toed gecko doesn’t actually go into brumation, it just slows down its activity during the coldest periods. Most adults are inactive and frequently only juveniles are active at air and substrate temperatures below 10° or 5°C (author pers. obs.). Mating takes place at the end of the winter and is facilitated by winter aggregations, which can consist of one or more dozens of individuals, depending of the size and the quality of the retreat site (Delaugerre, 2003). Most females are gravid from April to mid-July. Gravid females are the exception in autumn. Some may retain their late clutch throughout the winter. As in the case of a female with fully developed eggs, observed during nocturnal activity at the beginning of May 2021 in an alpine location of central Corsica (elevation of 1450 m asl, air temperature 3.7°C) author pers. obs. The first hatchings take place at the beginning of August and most occur from mid-August to early October. The 100% of juveniles below 22 mm SVL observed on the Corsican island of Gargalu (Table S2) in the early days of April were most likely not hatchlings, but juveniles around 6 months old with very low growth, as Gargalu are dwarf sized (Delaugerre and Cheylan, 1992). This dataset confirms and refines what was previously published from the single population of Port-Cros. Most females are gravid in April; oviposition lasts from mid-May to the end of July and hatchlings appear from mid-August to early October. Growth appears to be constant for the first two years, then slows in the third year, when functional sexual maturity is reached in both sexes (Salvidio and Delaugerre, 2003). The timing of the reproduction is indeed seasonal, but it is spread out over several weeks. With the comprehensive data set provided here, further studies and meta-analysis will be able to investigate a change in the timing of reproduction in relation to variations in weather conditions (Marco and Pérez-Mellado, 1998) even night-time temperatures (Rutschmann et al., 2016), clinal variation or a seasonal shift induced by climate change (Parmesan et al., 1999; Henle et al., 2008; Beaumont et al., 2015; Ljungström, Wapstra, and Olsson, 2015).
Acknowledgments
Most of the fieldwork was carried out in protected areas, national parks and nature reserves in Tunisia, Sardinia, Corsica and Provence, with the help of the managers. Many people (too many to mention) helped with night sessions on remote islands. I would like to thank all of these people and organisations, and Julien Renet and Yannick Turpin for giving us permission to use their photographs. A special dedication to my friend and instigator Pietro Lo Cascio, to whom I sent a sample of the Euleptes phylogeny data set; his reply was “… I see a lot of interesting data, why didn’t you think of publishing it?”
Supplementary material
Supplementary material associated with this article can be found at: https://oaj.fupress.net/index.php/ah/article/view/17238/14577.
References
Beaumont, L., Hartenthaler, T., Keatley, M., Chambers, L. (2015): Shifting time: Recent changes to the phenology of Australian species. Clim. Res. 63: 203-214.
Brown, G.P., Shine, R. (2006): Why do most tropical animals reproduce seasonaly? Testing hypotheses on an Australian snake. Ecology 87: 133-143.
Chukwuka, C.O., Monks, J.M., Cree, A. (2023): Extreme tolerance for nocturnal emergence at low body temperatures in a high-latitude lizard: Implications for future climate warming. Conserv. Physiol. 11: coac082.
Delaugerre, M. (1981): Sur l’histoire naturelle de Phyllodactylus europaeus Gené, (Gekkonidae, Sauria, Reptiles). Port-Cros: Étude d’une population naturelle. Trav. Sci. Parc Nation Port-Cros 1980 6: 147-175.
Delaugerre, M. (1984): Sur l’écologie thermique des geckos Phyllodactylus europaeus, Hemidactylus turcicus et Tarentola mauritanica: Rythmes d’activité, températures et activité, répartition altitudinale. Trav. Sci. Parc Nat. Régional Corse 3: 96-121.
Delaugerre, M., Cheylan, M. (1992): Atlas de répartition des batraciens et reptiles de Corse. Ajaccio Parc naturel régional de Corse, Ecole pratique des hautes études.
Delaugerre, M.J. (2003): Synthèse et mise à jour des carnets de terrains des recherches conduites entre 1975 et 1985. Hyères, Parc National de Port-Cros.
Delaugerre, M.J., Corti, C. (2020): Tiny but “strong”: The European Leaf-toed gecko, Euleptes europaea, a terrestrial vertebrate able to survive on tiny islets. Isr. J. Ecol. Evol. 66: 223-230.
Delaugerre, M.-J., Sacchi, R., Biaggini, M., Lo Cascio, P., Ouni, R., Corti, C. (2019): Coping with aliens: How a native gecko manages to persist on Mediterranean islands despite the Black rat? Acta Herpetol. 14: 89-100.
Deso, G., Priol, P., Reynier, T., Renet, J. (2024): High occupancy of European leaf-toed gecko in two island stands of Eucalyptus sp.: Tree selection, habitat effect, and syntopy with other gecko species. Herpetol. Conserv. Biol. 19: 263-274.
Forrest, J.R. (2016): Complex responses of insect phenology to climate change. Curr. Opin. Insect Sci. 17: 49-54.
Henle, K., Dick, D., Harpke, A., Kühn, I., Schweiger, O., Settele, J. (2008): Climate change impacts on European amphibians and reptiles. In: Biodiversity and Climate Change: Reports and Guidance Developed under the Bern Convention Council of Europe Publishing, pp. 225-305. Strasbourg, France, Council of Europe.
Ibáñez, I., Primack, R.B., Miller-Rushing, A.J., Ellwood, E., Higuchi, H., Lee, S.D., Kobori, H., Silander, J.A. (2010): Forecasting phenology under global warming. Philos. Trans. R. Soc. B Biol. Sci. 365: 3247-3260.
Knoepffler, L.P. (1973): Le Phyllodactyle d’Europe, une intéressante relique de la faune insulaire méditerranéenne. Ann. Mus. Hist. Nat. Nice 1: 87-91.
Ljungström, G., Wapstra, E., Olsson, M. (2015): Sand lizard (Lacerta agilis) phenology in a warming world. BMC Evol. Biol. 15: 206.
Marco, A., Pérez-Mellado, V. (1998): Influence of clutch date on egg and hatchling sizes in the annual clutch of Lacerta schreiberi (Sauria, Lacertidae). Copeia: 145-150.
Mourgue, M. (1910): Étude sur le Phyllodactyle d’Europe (Phyllodactylus europaeus Gené). Feuille Jeun Nat. 40: 57-61.
Parmesan, C., Ryrholm, N., Stefanescu, C., Hill, J.K., Thomas, C.D., Descimon, H., Huntley, B., Kaila, L., Kullberg, J., Tammaru, T., Tennent, W.J., Thomas, J.A., Warren, M. (1999): Poleward shifts in geographical ranges of butterfly species associated with regional warming. Nature 399: 579-583.
Pianka, E.R., Vitt, L.J. (2003): Lizards windows to the evolution of diversity. Univ. California Press.
Righi, A.F., Nascimento, L.B., Galdino, C.A.B. (2012): Seasonal reproduction in the rock gecko Phyllopezus pollicaris from a rock field habitat in Southeastern Brazil. J. Herpetol. 46: 632-636.
Rutschmann, A., Miles, D.B., Le Galliard, J., Richard, M., Moulherat, S., Sinervo, B., Clobert, J. (2016): Climate and habitat interact to shape the thermal reaction norms of breeding phenology across lizard populations. J. Anim. Ecol. 85: 457-466.
Saint-Girons, H.H., Saint-Girons, M.-C. (1956): Cycle d’activité et thermorégulation chez les reptiles (lézards et serpents). Vie Milieu 7: 133-226.
Salvi, D., Berrilli, E., Bruni, G., Garzia, M., Gomes, V., Radi, G., Delaugerre, M.-J. (2023): The secret life of a rock-dweller: Arboreal acrobatics observed in the European leaf-toed gecko Euleptes europaea. Herpetozoa 36: 135-141.
Salvidio, S., Delaugerre, M. (2003): Population dynamics of the European leaf-toed gecko (Euleptes europaea) in NW Italy: Implications for conservation. Herpetol. J. 13: 81-88.
Salvidio, S., Lanza, B., Delaugerre, M.J. (2010): Euleptes europaea (Gené, 1839). In: Fauna d’Italia, p. 869. Milano, Edizioni Calderini de Il Sole 24 ORE.
Van Dyke, J.U. (2015): Cues for reproduction in squamate reptiles. In: Reproductive biology and phylogeny of Lizards and Tuatara, pp. 109-143. Rheubert, J.L., Siegel, D.S., Trauth, S.E., Eds, CRC Press, Boca Raton, Florida.
Vitt, L.J. (1986): Reproductive tactics of sympatric gekkonid lizards with a comment on the evolutionary and ecological consequences of invariant clutch size. Copeia 773-786.